The DNA Polymerase Machine in DNA Replication
Having discovered the structure and base-pairing rules of the DNA double helix, famous scientists James Watson and Francis Crick published an article in which they commented, “it has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material”(Campbell 2005). After this discovery gained momentum, scientists set out to discover what this mechanism for DNA replication could be. Initially, there was some debate about how the DNA replicated and which model it followed. The most widely accepted was the semi-conservative model in which each complementary strand of DNA acts as a template for the synthesis of a new strand, thus creating two new double helices each with one old and one new strand. Accomplishing this task requires the use of a complex copying machine composed many smaller machines.
The DNA replication machine begins work at certain points along the double helix called origins of replication. At these points, the enzyme helicase attaches to the DNA molecule and begins unwinding the double helix and breaking the hydrogen bonds between the two strands. Helicase is actually an active molecule that moves along the double helix and directly destabilizes the DNA strands (Betterton and Julicher 2005). This unwinding causes increased strain on the portion of the helix ahead of the replication fork, but this strain is alleviated by the enzyme topoisomerase. This enzyme wraps around the helix and makes a cut in one strand. While holding on to the damaged portion, topoisomerase allows the DNA to spin freely to correct overwinding or underwinding. It then reconnects the cut strand and the helix is relieved of stress (Goodsell 2006).
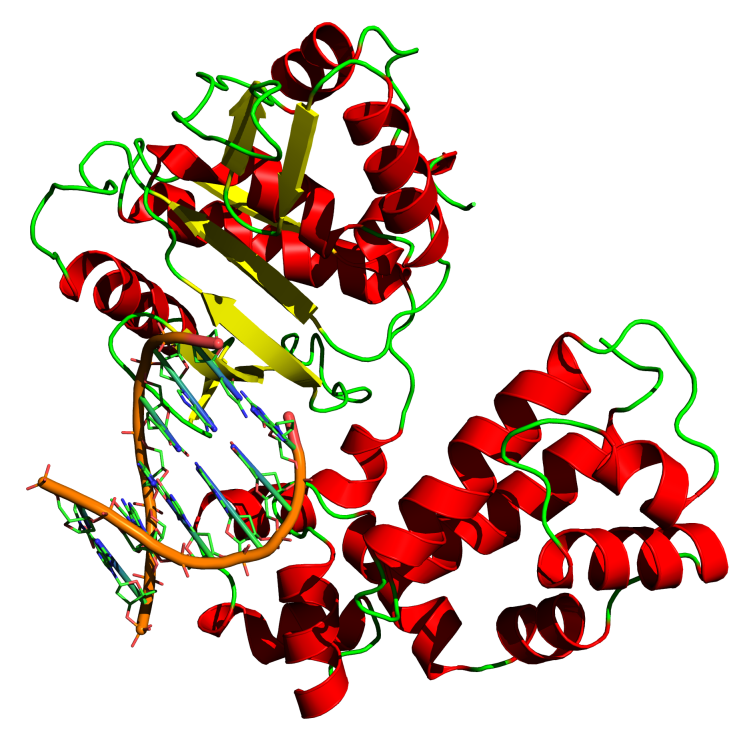
After helicase separates the strands, single-stranded binding protein then attaches to the strands and keeps them stable and separated until they are replicated. However, before new nucleotides can be added to the DNA strands, they must be primed. The enzyme primase creates an RNA strand that temporarily binds to the nucleotides at the origin of replication, creating a primer for the addition of new DNA nucleotides. Each of these primers is between five and ten nucleotides long (Campbell 2005). Once the DNA is unwound and primed, it is ready to enter the main part of the DNA replication machine, DNA polymerase III. Polymerase is shaped sort of like a hand, with the DNA helix being fed across the palm. There are three active sites on the polymerase molecule. The polymerase site adds nucleotides, the 3’-5’ exonuclease site proofreads new additions, and the 5’ exonuclease site removes RNA primers (Goodsell 2000). Starting with the primer on the 3’ end of the DNA strand, DNA polymerase III adds complementary nucleotides one at a time to the strand as it unwinds. Polymerase is an extremely accurate enzyme that proofreads as it copies, making less than one mistake for every billion bases added(Goodsell 2000). The nucleotides are added in the form of nucleoside triphosphates. As they are added to the growing DNA strand, two phosphates are lost as a pyrophosphate group which will power the polymerization reaction.
Figure 1. DNA Polymerase III
(Pelletier and Sawaya 1996)
Each strand of the double helix has a 3’ end and a 5’ end, and the two strands in the helix lie anti-parallel to each other. Similarly, the complementary daughter strands must also be anti-parallel to the parent strands. Because DNA replication can only occur in the 5’ to 3’ direction, two separate methods for replication are needed. For the strand that is copied in the direction toward the replication fork, DNA polymerase can add nucleotides continuously as the strand unwinds. This strand is called the leading strand. The strand that is being synthesized in the direction away from the replication fork, however, cannot be created continuously. Because this so called lagging strand is synthesized in the 5’ to 3’ direction, DNA polymerase III must start adding nucleotides near the replication fork and continue away from it. This process creates short segments of DNA called an Okazaki fragments, so named for the scientist who discovered them (Campbell 2005). In eukaryotes, these fragments are usually between 100 and 200 nucleotides long. As helicase unwinds the DNA, an Okazaki fragment is started and continues until it reaches the start of the last Okazaki fragment. Thus formation of the lagging strand is segmented and slightly delayed as compared with the leading strand. For the start of each Okazaki fragment, primase must add a new primer to the DNA strand. After the segments are formed, DNA polymerase I replaces the RNA primer with DNA. The joining of the Okazaki fragments employs the use of yet another enzyme, DNA ligase, which joins the sugar-phosphate backbones of the fragments. DNA ligase repairs these breaks in the DNA strands using ATP as a cofactor. Ligase is also helpful repairing DNA that has been broken by mistake such as a result of gamma radiation, or broken on purpose such as in meiosis (Goodsell 2004).
Figure 2. Replication process overview
(Campbell 2005)
All of these protein enzymes that contribute to DNA replication are actually joined to form one DNA replication machine. These proteins work best when they are in contact with each other, serving as integral parts of a single machine. Studies have shown that this DNA machine is stationary and that the polymerase actually feeds the parent DNA through the machine and expel the daughter strands. Several of theses machines may anchor close together in the nucleus at ‘replication sites’ to form replication ‘factories’ (Frouin, Montecucco et al. 2003). There is also evidence that the lagging strand is looped through the machine so that DNA polymerase III can easily transition between Okazaki fragments. Several protein-protein interactions facilitate the replication process, requiring parts of the machine to be close to one another. Helicase, for example, works more rapidly when it is in contact with primase (Campbell 2005).
Figure 3. Replication factory containing several replication machines
(Frouin, Montecucco et al. 2003)
There is, however, one downfall of the DNA replication machine. Because it must synthesize DNA in the 5’ to 3’ direction, the machine cannot complete the 5’ end of the new DNA strands. When the primer from an Okazaki fragment is removed, it cannot be replaced by DNA because polymerase I can only add DNA to the 3’ end. Therefore, the DNA strand gets shorter each time it is replicated. To prevent the eroding of important genetic material as a result of replication, DNA strands end with nucleotide sequences called telomeres. These telomeres do not contain genes but repeated patterns of nucleotide sequences, which in the case of humans is TTAGGG and is repeated anywhere from 100 to 1,000 times in a telomere. By having these non-information carrying sequences on the DNA molecules, the genetic information is protected from erosion because the telomere is eroded instead. For DNA in cells that must persist for many generations without being eroded, such as germ cells, the enzyme telomerase helps restore the length of the telomere. The DNA saving properties of polymerase provide hope for anti-cancer treatments that target the telomeres of cancer cells (Campbell 2005).
Though this DNA copying machine is quite complex, it is probably the most accurate machine in the world. It is also one of the most abundant. Organisms ranging from bacteria to humans use similar copying machines to replicate their DNA. While the major structures and functions of this machine have been discovered, there is still much to learn about it. Having harnessed the power of this machine, scientists have already been able to use it in processes such as polymerase chain reaction and cloning. By continuing to study this machine, the future will undoubtedly hold more uses for its speed and accuracy in copying DNA.
Works Cited
Betterton, M. D. and F. Julicher (2005). "Opening of nucleic-acid double strands by helicases: Active versus passive opening (vol 71, pg 11904, 2005)." Physical Review E 72(2): -.
Campbell, N. (2005). The Molcular Basis of Inheritance. Biology. Wilbur. San Francisco, Benjamen Cummings: 293-307.
Frouin, I., A. Montecucco, et al. (2003). "DNA replication: a complex matter (vol 4, pg 666, 2003)." Embo Reports 4(8): 766-766.
Goodsell, D. (2000). "DNA Polymerase." Molecule of the Month Retrieved February 23, 2008, from http://www.rcsb.org/pdb/static.do?p=education_discussion/molecule_of_the_month/index.html.
Goodsell, D. (2004). "DNA Ligase." Molecule of the Month Retrieved February 23, 2008, from http://www.rcsb.org/pdb/static.do?p=education_discussion/molecule_of_the_month/pdb55_1.html.
Goodsell, D. (2006). "Toposisomerases." Molecule of the Month Retrieved February 24, 2008, from http://www.rcsb.org/pdb/static.do?p=education_discussion/molecule_of_the_month/pdb73_1.html.
Pelletier, H. and M. R. Sawaya (1996). "Characterization of the metal ion binding helix-hairpin-helix motifs in human DNA polymerase beta by X-ray structural analysis." Biochemistry 35(39): 12778-12787.
{kind=link}
Subscribe to:
Post Comments (Atom)
No comments:
Post a Comment